- 品牌
- 上海云序生物
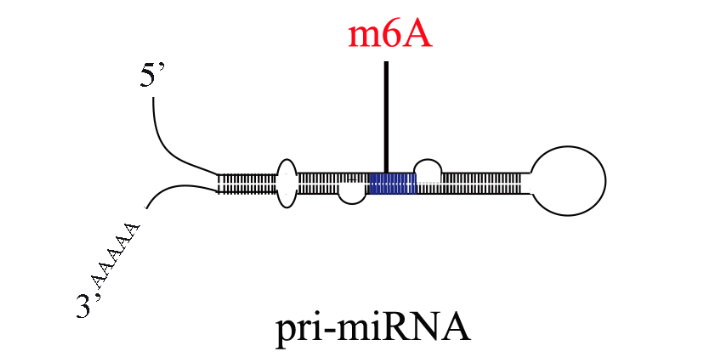
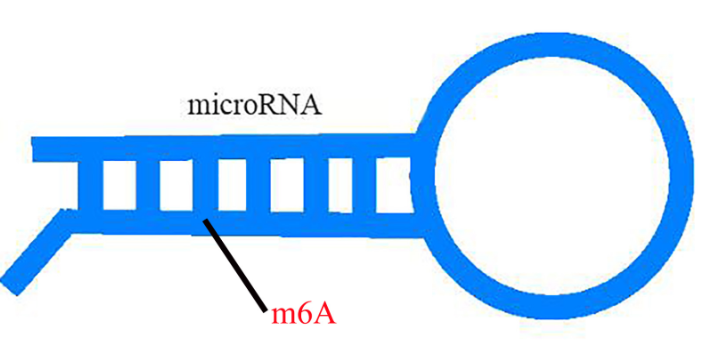
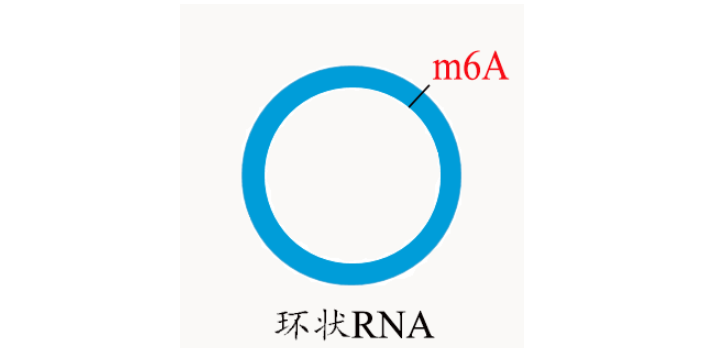
m6A甲基化已经被证明与植物对病原体的抗性有关。然而,小麦(Triticum aestivum) 全转录组m6A谱及其在小麦抗小麦黄花叶病毒(WYMV)中的潜在生物学功能尚未见报道。这项研究是shou次鉴定两个不同抗WYMV小麦品种的转录组m6A修饰谱。通过对m6A-MeRIP-seq数据的分析,作者在WYMV感ran的抗病小麦品种(WRV)和WYMV感ran的敏感小麦品种(WSV)中鉴定出25752个共有m6A峰和30582个共有m6A基因,这些峰主要富集在编码序列3‘UTR区和终止密码子中。GO分析和RNA测序数据显示,m6A和mRNA水平均发生明显变化的基因与植物防御反应有关。而随后Cell Research也发表文章,证实环状RNA也会被m6A甲基化修饰。上海甲基化筛选
RNA 修饰在基因表达调控中扮演着非常重要的角色。m6A 修饰就是真核生物 mRNA 中较常见存在的一种修饰形式,得到了常见的关注和研究。除此之外,还存在着一种分子结构上非常相似的 RNA 修饰形式:m6Am——在 m6A 修饰的基础上,同一个腺苷酸残基的核糖的 2’ 羟基也被甲基化,产生 2’ 甲氧基结构(2’-O-CH3)。与 m6A 的常见分布所不同的是,m6Am 修饰的分布主要局限于 mRNA 5’ 帽子结构下游的较早核苷酸残基上发生。该位点的 m6Am 修饰在进化上高度保守,无论是在斑马鱼、小鼠还是人类细胞中均有发现。组蛋白甲基化过程RNA m6A甲基化是RNA较关键的内部修饰之一。
m7G RNA甲基化是在甲基化转移酶的作用下,使RNA鸟嘌呤(G)的第七位N上加上甲基的一种修饰( N7-methylguanosine,m7G)。研究表明,m7G RNA甲基化修饰存在于各类分子中,包括:mRNA 5’帽子结构、mRNA内部、pri-miRNA、转运RNA(tRNA)和核糖体RNA(rRNA)。m7G RNA甲基化修饰能够调节mRNA的转录、miRNA的生物合成和生物学功能、tRNA稳定性、18S rRNA的核内加工及成熟。m7G RNA甲基化修饰作为一类新型RNA甲基化,近两年高影响因子文章不断,是继m6A修饰之后的又一表观转录组学热点。
ac4C RNA乙酰化是在RNA ac4C修饰酶的作用下,使N4位乙酰胞嘧啶发生乙酰化的一种保守的化学修饰(N4-acetylcytidine)。早期研究发现该修饰存在于真核生物中丝氨酸及亮氨酸tRNA和18S rRNA上,导致Watson-Crick碱基热稳定性增加,调控蛋白合成中的编码准确性;近期研究发现ac4C分布在人类转录组中,大多数位点出现在编码序列(CDS)内,并且通过改善的mRNA稳定性和翻译促进靶基因表达。目前,ac4C RNA乙酰化修饰作为一类新型RNA修饰,是继m6A修饰之后的又一表观转录组学热点和“超级潜力股”!ctDNA 的甲基化检测,同时结合了 ctDNA 测序于 DNA 甲基化测序的优势。
案例3:拟南芥m5C RNA甲基化谱及TRM4B酶对根的影响 原文:Transcriptome-Wide Mapping of RNA 5-Methylcytosine in Arabidopsis mRNAs and Noncoding RNAs 期刊:The Plant Cell 影响因子:8.23 与上篇不同,本研究团队采用m5C RNA甲基化测序研究拟南芥中m5C甲基化谱,在种子,幼芽,根中发现了1000多个特异性的位点。敲低RNA m5C甲基转移酶TRM4B,造成tRNA稳定性的降低。研究人员还证实TRM4B突变体的初级根比野生型更短,同时对氧化的应激反应更敏感。m7G RNA甲基化修饰作为一类新型RNA甲基化。850KDNA甲基化表达
miRNA的生物合成和生物学功能、tRNA稳定性、18S rRNA的核内加工及成熟。上海甲基化筛选
云序生物对O8G RNA氧化化检测手段为O8G-IP-Seq技术,技术原理如下: 将O8G特异性抗体与被随机打断的RNA的片段进行共孵育,抓取有O8G修饰的片段进行测序;同时需要平行测序一个对照(Input)样本,对照样本只含有打断的RNA的片段,并不添加O8G特异性抗体与其共孵育。对照样本用于消除非特异性抓取的O8G片段的背景。对比免疫共沉淀(IP)样本和对照样本(Input)中的序列片段,将O8G氧化修饰位点定位到转录组上,并根据RNA-seq数据,计算样本中O8G氧化程度及对RNA表达水平的影响。上海甲基化筛选
- 吉林DNA甲基化 2023-08-07
- 河北miRNA甲基化 2023-08-07
- 内蒙古tRNA甲基化 2023-08-07
- 宁夏超微量RNA甲基化 2023-08-07
- 甲基化表达 2023-08-07
- 云南甲基化研究 2023-08-07
- 外泌体甲基化过程 2023-08-07
- 四川pri-miRNA甲基化 2023-08-07
- 辽宁microRNA甲基化 2023-08-06
- DNA甲基化调控 2023-08-06
- 陕西microRNA甲基化 2023-08-06
- 西藏甲基化测序 2023-08-06