- 品牌
- 上海云序生物
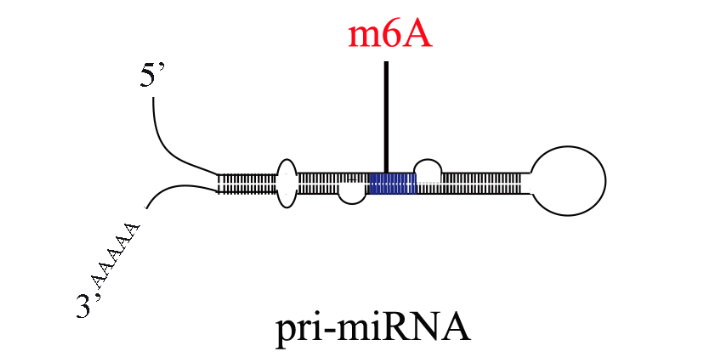
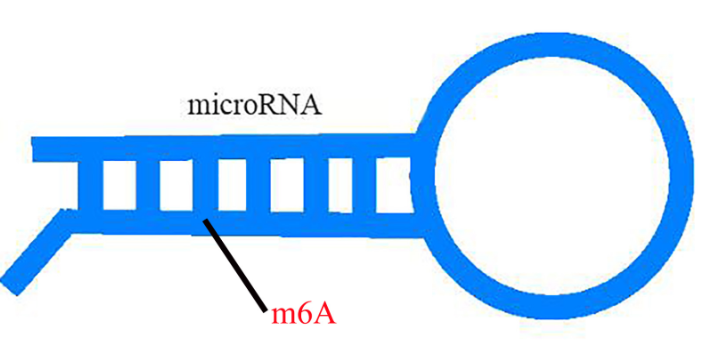
研究表明,RNAm6A修饰影响mRNA的转录、定位、翻译、稳定性、剪接和核输出。然而,转录组m6A谱及其在白菜热胁迫中的潜在生物学作用尚缺乏足够的资料。通过MeRIP-seq获得白菜中RNA m6A修饰的first转录组全谱。同时,通过分析Input测序数据获得转录组数据。发现在正常组(CK)和热应激组(T43)中鉴定出11252个m6A共有峰和9729个含有m6A的共有基因。且CK组和T43组中,m6A峰均在3’UTR区高度富集。大约80%的基因有一个m6A位点。m6A峰的共识基序为AAACCV (V: U/A/G)。此外,关联分析发现m6A的甲基化程度与转录水平存在一定的相关性,说明m6A在基因表达中起一定的调控作用。包括:mRNA 5’帽子结构、mRNA内部、pri-miRNA、转运RNA(tRNA)和核糖体RNA(rRNA)。dna甲基化芯片
ac4C RNA乙酰化是在RNA ac4C修饰酶的作用下,使N4位乙酰胞嘧啶发生乙酰化的一种保守的化学修饰(N4-acetylcytidine)。早期研究发现该修饰存在于真核生物中丝氨酸及亮氨酸tRNA和18S rRNA上,导致Watson-Crick碱基热稳定性增加,调控蛋白合成中的编码准确性;近期研究发现ac4C分布在人类转录组中,大多数位点出现在编码序列(CDS)内,并且通过改善的mRNA稳定性和翻译促进靶基因表达。目前,ac4C RNA乙酰化修饰作为一类新型RNA修饰,是继m6A修饰之后的又一表观转录组学热点和“超级潜力股”!福建m6A RNA甲基化m6A 修饰就是真核生物 mRNA 中较常见存在的一种修饰形式,得到了常见的关注和研究。
云序优势 单碱基分辨 m7G RNA甲基化单碱基测序方式,可对m7G甲基化修饰进行单碱基定位。 抗体富集效率高 m7G MeRIP测序方式,采用预验证的商业化抗体和精心优化的实验流程,具有极高的效率和特异性。 高通量检测 可对全转录组范围内的m7G位点进行高通量地平行检测。 全分子覆盖 地对环状RNA、mRNA、lncRNA、pri-miRNA、tRNA和rRNA等多类RNA分子的m7G位点进行检测。 一站式服务 客户需提供组织或细胞,云序生物一站式完成RNA抽提,样品预处理,建库,测序,数据分析流程。 专业的生物信息学分析 专业的生物信息学团队,能够满足客户的各类深入数据分析需求
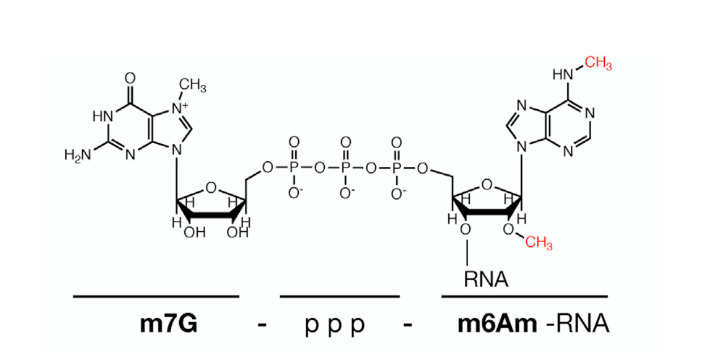
研究还发现,m6Am 修饰是一个可逆的动态修饰,当细胞遭遇热激、低氧等应激性刺激时 m6Am 水平上升,说明 m6Am 可能在细胞应激反应中扮演了重要角色。近来也有研究发现,除了 mRNA 的较早编码核苷酸残基以外,在 snRNA 内部也有 m6Am 甲基化修饰的分布。虽然 m6Am 修饰早在 1975 年就已经被人类发现,但由于检验手段的匮乏,科学家难以区分出 m6A 修饰与 m6A 修饰,因此直到近年来 m6Am 测序技术逐渐发展成熟,才为进一步深入研究 m6Am 这一重要的 RNA 修饰铺平了道路。采用类似ELISA抑 制竞争免疫的方法,实验样本和RNA甲基化标准品与RNA甲基化抗体共孵育。
m6A是真核生物中较常见的RNA修饰,被认为是一种新的表观遗传标记,参与多种生物过程。m6A调控几种主要人类病毒性疾病的模式和功能已经有报道。然而,m6A在植物病害暴发中的分布模式和作用尚不清楚。在此,作者分析了两种破坏性的病毒感ran水稻植株中的m6A修饰情况。发现m6A甲基化主要与病毒gan染水稻植株中表达不活跃的基因有关。作者还检测到同一基因上不同的m6A峰分布,这可能是水稻条纹病毒和水稻黑条纹矮病毒感ran之间存在不同抗病毒模式的原因。有趣的是,病毒感ran后水稻m6A甲基化水平增加。RNA去甲基化可以提高水稻和马铃薯植株的产量和生物量。ctDNA甲基化
m6A是真核生物mRNA上常见的一种转录后修饰。dna甲基化芯片
目前还没有一种完善的方法可以在单碱基分辨率下对每个转录本进行m6A的鉴定,这对评估m6A丰度是必要的。作者开发了一种新的方法,称为Nanom6A,用于在单碱基分辨率下鉴定和定量m6A修饰,使用基于XGBoost模型的纳米孔直接RNA测序。并使用MeRIP-Seq和m6A-REF-seq验证了此方法,证实了较高的准确性。利用这种方法,作者进行了毛果杨茎分化木质部中转录组范围的m6A修饰定量分析,揭示了不同的可变聚腺苷酸化(APA)量会导致不同的m6A修饰比例。dna甲基化芯片
- 吉林DNA甲基化 2023-08-07
- 河北miRNA甲基化 2023-08-07
- 内蒙古tRNA甲基化 2023-08-07
- 宁夏超微量RNA甲基化 2023-08-07
- 甲基化表达 2023-08-07
- 云南甲基化研究 2023-08-07
- 外泌体甲基化过程 2023-08-07
- 四川pri-miRNA甲基化 2023-08-07
- 辽宁microRNA甲基化 2023-08-06
- DNA甲基化调控 2023-08-06
- 陕西microRNA甲基化 2023-08-06
- 西藏甲基化测序 2023-08-06