- 产地
- 苏州
- 品牌
- 细胞外基质胶
- 型号
- 齐全
- 是否定制
- 是
细胞外基质与医学:恶性部位的发生、发展、侵袭和转移常常伴有细胞外基质(extracellularmatrix,ECM)及其细胞表面受体表达的变化。正常肝细胞没有基膜,也不表达层粘连蛋白(laminin,LN)的特异性整合素族受体α6β1;而在肝细胞(humanhepatocellularcarcinoma,HCC)组织中,LN和α6β1不仅表达水平升高,呈明显的共分布,而且其高水平表达与肝患者的预后呈负相关,提示HCC细胞可能通过α6β1受体接受来自LN的信号,从而对肝细胞的侵袭行为起着不可忽视的作用。肝的发病过程中往往早期就出现门静脉侵袭、肝内转移以及肝外肺脏和骨组织的转移,肝的侵袭、转移和术后复发是影响患者预后的主要因素。基质金属蛋白酶(matrixmeta-lloproteinases,MMPs)对ECM的降解是部位细胞侵袭和转移的关键环节之一,多种恶性部位都伴有MMPs分泌水平和活性的增高。细胞通过mTORC1感知营养素可用性并相应地在分解代谢和合成代谢状态之间转换来协调全身和细胞代谢。上海天津细胞外基质胶

功能:由于其不同的性质和组成,细胞外基质可以发挥许多功能,例如提供支持、将组织相互隔离以及调节细胞间的通讯。细胞外基质调节细胞的动态行为。此外,它能隔离多种细胞生长因子,并充当它们的局部储存库。生理条件的变化会引发蛋白酶活性,从而导致这类物质在局部释放。这允许快速和局部生长因子介导的细胞功能,而无需从头合成。这种效应已经在一项模型和理论研究中进行了探索,其中血管内皮生长因子C(VEGFC)、基质金属蛋白酶2(MMP2)和胶原蛋白I被用作研究例子。南昌正规细胞外基质胶厂家批发价细胞外基质将细胞连接在一起,形成组织、部位,而是含有大量信号分子,积极参与控制细胞的生长。

免疫系统和细胞外基质之间的串扰:被囊动物是脊索动物中的尾索动物,成年海鞘包被着由被膜组成的细胞外基质。伤口愈合过程中,被膜基质被免疫细胞重塑,如脱粒细胞。细菌与海鞘之间的相互联系已有报道,可能涉及分泌活性产物,如克菌蛋白与吞噬细胞一起作为先天免疫系统的一部分。在棘皮动物中,造血组织被描述为分泌体腔细胞的体腔上皮,这些是伤口愈合的重要调节剂,因为它们迁移到伤口部位形成血块,并在ECM的调节中发挥作用。在海参中体腔细胞可能是免疫反应和伤口修复的重要调节器。
细胞外基质成分居然能调节葡萄糖代谢过程:细胞具有不同的机制以感知和响应外在代谢信号。例如,细胞通过mTORC1感知营养素可用性并相应地在分解代谢和合成代谢状态之间转换来协调全身和细胞代谢;生长因子,和细胞因子可以将代谢信号传递给相邻细胞和远端组织,作为更普遍的生物反应的一部分。统一效应将细胞的行为和代谢与组织和生物体的需求结合起来。虽然ECM重塑和升高的糖酵解在多种生物学背景下是一致的,但这些过程之间的机制联系尚未确定。控制细胞的分化细胞通过与特定的细胞外基质成分作用而发生分化。细胞外基质:在生物学中,细胞外基质是细胞外大分子(如胶原蛋白、酶和糖蛋白)组成的三维网络。

这种转化细胞接种入正常机体,常能长成块,并侵润正常组织,发生普遍转移。如何理解细胞外基质影响细胞的粘附过程:1.决定细胞的形状体外实验证明,各种细胞脱离了细胞外基质呈单个游离状态时多呈球形.同一种细胞在不同的细胞外基质上粘附时可表现出完全不同的形状.上皮细胞粘附于基膜上才能显现出其极性.细胞外基质决定细胞的形状这一作用是通过其受体影响细胞骨架的组装而实现的.不同细胞具有不同的细胞外基质,介导的细胞骨架组装的状况不同,从而表现出不同的形状.2.控制细胞的分化细胞通过与特定的细胞外基质成分作用而发生分化.例如,成肌细胞在纤粘连蛋白上增殖并保持未分化的表型;而在层粘连蛋白上则停止增殖,进行分化,融合为肌管。细胞外基质生理学功能:趋硬性:硬度和弹性也引导细胞迁移,这一过程被称为趋硬性。贵阳细胞外基质胶报价
细胞外基质中不同的机械特性对细胞行为和基因表达都有影响。上海天津细胞外基质胶
细胞外基质的组分及组装形式由所产生的细胞决定,并与组织的特殊功能需要相适应。例如,角膜的细胞外基质为透明柔软的片层,肌腱的则坚韧如绳索。细胞外基质不光静态的发挥支持、连接、保水、保护等物理作用,而且动态的对细胞产生很全影响。细胞外基质的成分:构成细胞外基质的大分子种类繁多,可大致归纳为四大类:胶原、非胶原糖蛋白、氨基聚糖与蛋白聚糖、以及弹性蛋白。上皮组织、肌组织及脑与脊髓中的ECM含量较少,而结缔组织中ECM含量较高。上海天津细胞外基质胶
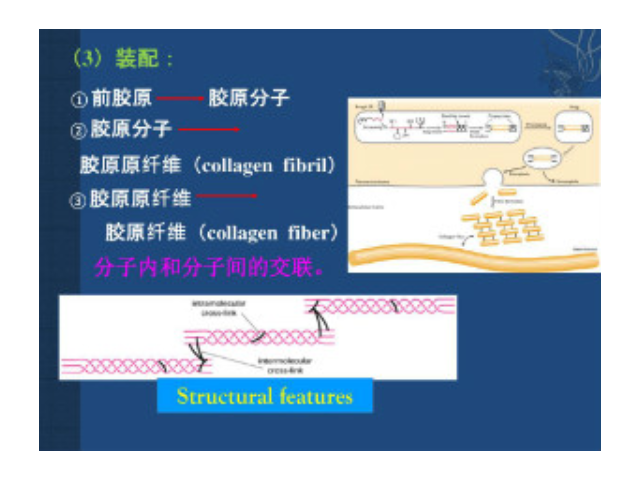
构成细胞外基质的大分子:原胶原共价交联后成为具有抗张强度的不溶性胶原。胚胎及新生儿的胶原因缺乏分子间的交联而易于抽提。随年龄增长,交联日益增多,皮肤、血管及各种组织变得僵硬,成为老化的一个重要特征。人α1(Ⅰ)链的基因含51个外显子,因而基因转录后的拼接十分复杂。翻译出的肽链称为前α链,其两端各具有一段不含Gly-X-Y序列的前肽。三条前α链的C端前肽借二硫键形成链间交联,使三条前α链“对齐”排列。然后从C端向N端形成三股螺旋结构。前肽部分则呈非螺旋卷曲。带有前肽的三股螺旋胶原分子称为前胶原(procollagen)。胶原变性后不能自然复性重新形成三股螺旋结构,原因是成熟胶原分子的肽链不含前肽...
- 金华北京细胞外基质胶 2026-02-03
- 上海天津细胞外基质胶 2026-02-03
- 青岛正规细胞外基质胶供应商 2026-02-03
- 芜湖正规细胞外基质胶厂家推荐 2026-02-03
- 成都细胞外基质胶厂家批发价 2026-02-03
- 太原细胞外基质胶销售厂家 2026-02-03
- 重庆正规细胞外基质胶厂家供应 2026-02-03
- 重庆细胞外基质胶推荐厂家 2026-02-03
- 成都正规细胞外基质胶厂家推荐 2026-02-03
- 唐山细胞外基质胶 2026-02-03



- 温州正规细胞外基质胶直销厂家 2026-02-02
- 细胞外基质胶生产厂家 2026-02-02
- 正规细胞外基质胶哪家好 2026-02-02
- 无锡正规细胞外基质胶直销价 2026-02-02
- 深圳正规细胞外基质胶价格 2026-02-02
- 珠海正规细胞外基质胶厂家供应 2026-02-02



- 芜湖正规细胞外基质胶厂家推荐 02-03
- 成都细胞外基质胶厂家批发价 02-03
- 太原细胞外基质胶销售厂家 02-03
- 重庆正规细胞外基质胶厂家供应 02-03
- 南昌RNA提取试剂厂家 02-03
- 苏州鼠尾胶原单价 02-03
- 重庆细胞外基质胶推荐厂家 02-03
- 成都正规细胞外基质胶厂家推荐 02-03
- 唐山细胞外基质胶 02-03
- 上海无血清细胞冻存液厂家批发价 02-03