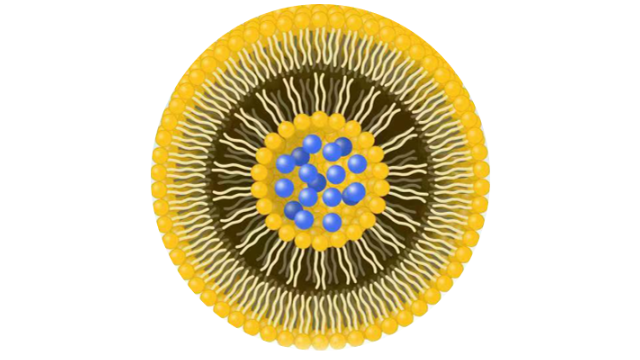
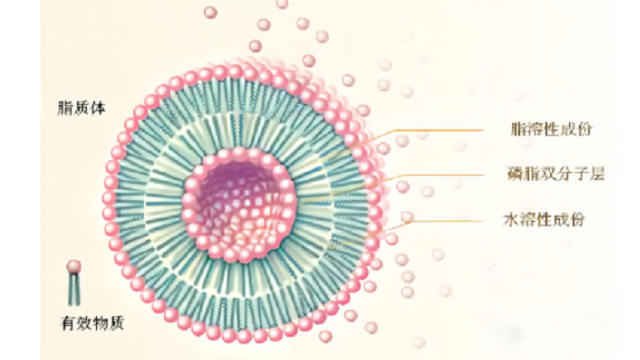
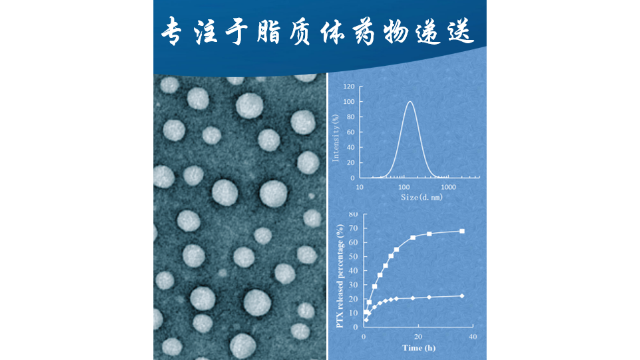
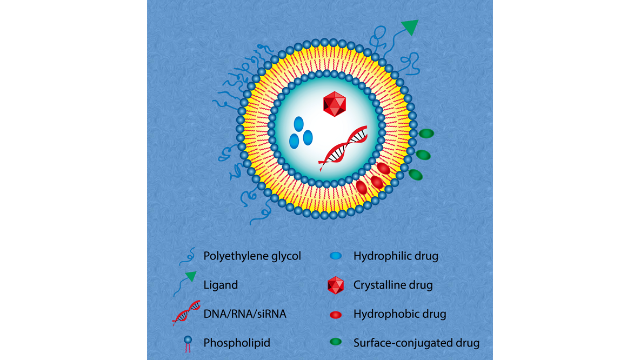
3脂质体中的相变温度
脂质体中的相变温度是指脂质双分子层中脂质分子从一个状态转变为另一个状态所需的温度。这个温度对于脂质体的性质和功能具有重要作用:1.药物释放控制:脂质体在体内可以通过温度变化来控制药物的释放。例如,如果脂质体的相变温度在人体温度范围内,那么在注射进体内后,脂质体可能会在特定温度下释放药物,这可以用于设计温敏***物输送系统。2.稳定性:相变温度也可以影响脂质体的稳定性。在相变温度以下,脂质体可能会形成固态结构,增加了其稳定性,而在相变温度以上,脂质体可能会转变为液态,导致结构松散和药物释放。3.生物相容性:脂质体的相变温度应当与生物体的温度相匹配,以确保脂质体的生物相容性。如果相变温度太高或太低,可能会对组织或细胞产生不良影响。负载药物的选择:相变温度也可能影响到可负载在脂质体中的药物类型。一些药物可能会干扰脂质体的相变温度,而另一些药物则可能受到相变温度的影响,导致在特定温度下释放。表明脂质体双分⼦层在体温中处于⽆序和药物“漏出”状态。综上所述,脂质体中的相变温度对于药物输送系统的设计和性能调控非常重要,可以影响药物的释放速率、稳定性和生物相容性。 脂质体中的相变温度是指脂质双分子层中脂质分子从一个状态转变为另一个状态所需的温度。江苏纳米脂质体载药

siRNA脂质体
RNA干扰(RNAi)途径允许siRNA和miRNAs负向调节蛋白表达。siRNA是21~23对核苷酸组成的双链RNA,可诱导同源靶mRNA沉默。为了发挥作用,双链siRNA分裂成两个单链RNA:乘客链和引导链。乘客链被argonaute-2蛋白降解,而引导链则被纳入RNAi诱导的沉默复合体中,该复合体结合与引导链互补的mRNA并将其切割。siRNA似乎具有***多种疾病的巨大潜力,因为它们可以很容易地下调各种靶mRNA,而不考虑它们的位置(即在细胞核或细胞质中),并且它们的特异性结合表明它们比传统化学药物诱导的副作用更少。作为一种新型的基于核酸的***策略,siRNA***与传统的化学药物相比具有许多优势。然而,为了促进基于siRNA的***方法的发展,必须克服一些挑战,包括需要识别适当的靶基因和开发优化的递送系统。许多研究人员试图利用阳离子脂质体提高siRNA的细胞递送和基因沉默效率。例如,由DC-6-14、DOPE和胆固醇组成的阳离子脂质体被用于递送萤火虫荧光素酶特异性的siRNA。当阳离子脂质体与siRNA持续剧烈搅拌混合时,转染效率提高,说明将siRNA加载到阳离子脂质体上的方法可以调节转染效率。siRNA脂丛的***应用因靶蛋白而异。 江苏纳米脂质体载药脂质体质量控制的重要性。
脂质体疫苗通常在已知疫苗中使用纯化抗原或减毒病原体作为免疫原。然而,长期的免疫反应可能不会由纯化抗原诱导,甚至有时根本不会诱导反应。另一方面,减毒疫苗可以在免疫的患者中产生应答。然而,递送包裹在脂质体内的抗原可诱导长期应答,这在某些抗原的直接免疫中没有观察到。研究表明,恶性细胞的细胞膜可以形成包封潜在抗原的脂质体。文献报道了包封在脂质体中的肽作为**疫苗的***应用能力。有研究评估了BLP25(一个含有合成人MUC1肽的25个氨基酸序列)作为**疫苗的能力。用二硬脂酰磷脂酰胆碱、胆固醇和二肉豆醇酰磷脂酰甘油(摩尔比3:1:25)中含有的单磷脂酰脂A(1%w/w)制备脂质体,然后与脂偶联和非偶联的MUC1肽结合。C57BL/6小鼠免疫分别采用肽相关脂质体、肽与无肽脂质体混合、脂肽单独免疫。结果表明,脂质体制剂对免疫应答有深远的影响。与物理相关的脂质体观察到强烈的免疫反应(抗原特异性t细胞细胞反应),而与肽混合的无肽脂质体或单独的脂肽则没有。体液免疫反应受到关联性质的***影响,这可以通过表面暴露的肽脂质体诱导muc1特异性抗体来证明。因此可以通过调整脂质体药物传递系统来诱导优先细胞反应这提出了一个假设即不同的脂质体配方刺激不同的免疫途径。
脂质体靶向递送中甘露糖配体修饰由于在巨噬细胞上发现了甘露糖受体,因此甘露糖已被用于修饰阳离子脂质体以供巨噬细胞递送。为了抑制由活化的巨噬细胞诱导的破骨细胞生成,将甘露糖基化阳离子脂质体与双链寡核苷酸NFkB诱饵络合。甘露糖阳离子脂质体/NFkB诱饵复合物有效诱导NFkB活化并抑制肿瘤坏死因子-a的产生。在另一项研究中,巨噬细胞靶向NFkB诱饵装载在甘露糖基化阳离子脂质体中,用于预防脂多糖诱导的肺部炎症。气管内给药后,甘露糖标记的阳离子脂质体/NFkB诱饵复合物***下调NFkB的表达,减少肿瘤坏死因子-a和白细胞介素-1b的释放。研究人员研究了茴香酰胺修饰的阳离子脂质体将寡核苷酸靶向递送至表达sigma受体的细胞的能力。剪接开关寡核苷酸(SSOs)是一种单链寡核苷酸,可与剪接位点或剪接增强子结合,阻断内源性剪接机制的通路,并产生成熟mRNA的替代版本。在肺转移小鼠模型中,全身给药装载Bcl-xSSO的茴香胺修饰阳离子脂质体可降低**生长。由于在巨噬细胞上发现了甘露糖受体, 因此甘露糖已被用于修饰阳离子脂质体以靶向巨噬细胞递送。

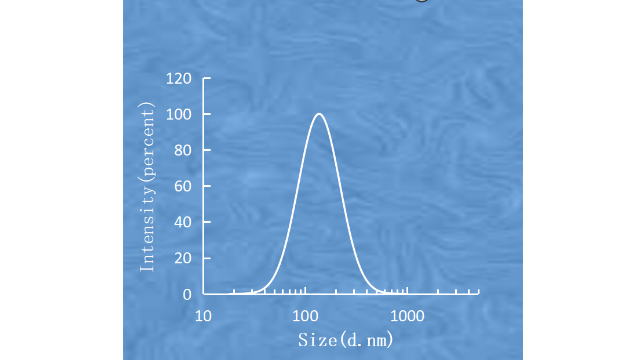
脂质体成功降低了绿色荧光蛋白(GFP)的表达,并在H4II-E和HepG2细胞中显示出较低的细胞毒性。在其他研究中,精氨酸衍生物N,N-distearyl-N-methyl-N-2-(N’-arginyl)aminoethylammoniumchloride被用于阳离子脂质体与胆固醇的配制。将这些离子脂质体与c-MycsiRNA络合,并静脉注射给B16F10黑色素瘤小鼠(1.2mg/kg,每天1次,连续3天),导致B16F10**对紫杉醇增敏。另一项研究建议使用精氨酸基DiLA2脂质作为载脂蛋白b特异性siRNA递送的阳离子脂质体组分。经小鼠静脉给药(ED50,0.1mg/kg)后,DiLA2和DOPE制备的阳离子脂质体显示出抑制肝脏载脂蛋白BmRNA表达的潜力。单次全身给药后,在给药后第2天观察到目标mRNA水平的比较大减少(约80%),并且目标mRNA的减少持续到给药后第9天。脂质体的粒径和粒径分布的检测。胰腺靶向脂质体载药对比剂
被动载药⽅法是在脂质体制备过程中对药物进⾏包封的方法。江苏纳米脂质体载药
两者都含有一种可电离的脂质,在低pH值下带正电荷(使RNA络合),在生理pH值下为中性(减少潜在的毒性作用并促进有效载荷释放)。它们还含有聚乙二醇化脂质,以减少血清蛋白的抗体结合(调理)和吞噬细胞的***,从而延长体循环。辉瑞公司的阳离子脂质:peg脂质:胆固醇:DSPC的摩尔比为(43:1.6:47:9.4),莫当纳疫苗的摩尔比为(50:1.5:38.5:10)。这些纳米颗粒直径为80 - 100纳米,每个脂质纳米颗粒含有大约100个mRNA分子。ALC-0315(辉瑞)和SM-102 (Moderna)这两种脂质都是叔胺,在低ph下质子化(因此带正电荷)。它们的碳氢链通过可生物降解的酯基连接,在mRNA传递后能够安全***。mRNA疫苗中使用的阳离子脂质含有支链烃链,这优化了非层状相的形成和mRNA的递送效率。peg -脂质均为PEG-2000偶联物。LNPs是在低pH (pH 4.0)条件下制备的,在这种条件下,可电离的脂质带正电,因此它很容易与mRNA形成复合物。微流控装置用于将水中含有mRNA的流与乙醇中含有脂质混合物的流混合。当快速混合时,这两种流的成分形成纳米颗粒,捕获带负电荷的mRNA。江苏纳米脂质体载药