- 品牌
- 上海云序生物
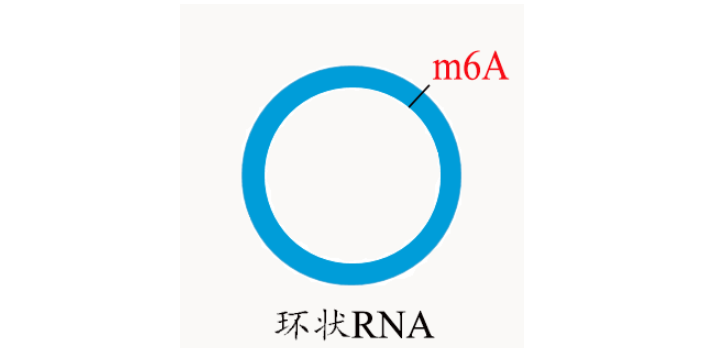
LncRNA m6Am-Exo-seq 测序服务 云序生物在国内shou批引入 m6Am-Exo-seq 测序服务,利用 5’ 核酸外切酶消除非目的性的 RNA 的片段上 m6A 修饰的信号干扰,选择性地获取 mRNA以及 LncRNA 的 5’ 帽子结构下游 m6Am 修饰富集区域的序列信息。接下来,我们对选择性获取到的 m6Am 修饰的 RNA 的片段进行反转录建库和高通量测序(测序结果将同时包含 mRNA 和 LncRNA)。通过后续的生物信息学分析,可以区分来自 mRNA 和 LncRNA 的测序片段,从而较全揭示 LncRNA和mRNA 的 m6Am 修饰位点,并推测其可能的生物学功能。 m6A除了分布在 mRNA 中,也出现在很多非编码 RNA中 ,如:环状RNA 、LncRNA等。北京m7G+m3C RNA甲基化
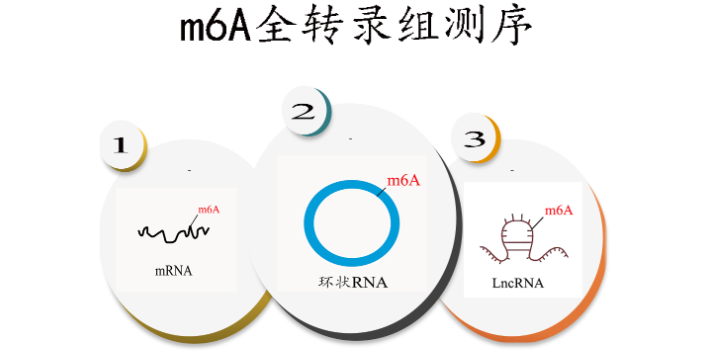
近年来,科学家们发现了一种可逆的RNA甲基化—m6A,即RNA分子腺嘌呤第6位氮原子上发生甲基化修饰(N6-methyladenosine,m6A)。研究发现,m6A是真核生物mRNA上常见的一种转录后修饰,m6A在细胞加速mRNA代谢和翻译,以及在细胞分化、胚胎发育和压力应答等过程中起重要作用。种种研究迹象表明m6A 存在于很多物种中,占到了 RNA甲基化修饰的80%。m6A除了分布在 mRNA 中,也出现在很多非编码 RNA中 ,如:环状RNA 、LncRNA等。Nature正刊发表文章,证实发生m6A RNA甲基化修饰可以调控mRNA稳定性。而随后Cell Research也发表文章,证实环状RNA也会被m6A甲基化修饰,并会促进环状RNA编码蛋白这一有趣的结论。北京m7G+m3C RNA甲基化m6A甲基化是番茄果实mRNA中普遍存在的修饰。
案例1:m5C RNA甲基化调控mRNA出核新机制 原文:5-methylcytosine promotes mRNA export — NSUN2 as the methyltransferase and ALYREF as an m5C reader 期刊:Cell Research 影响因子:15.60 中科院汪海林等研究团队揭示了m5C修饰在mRNA的分布图谱规律及其对调控mRNA出核作用新机制。研究人员建立了改进的RNA m5C单碱基分辨率高通量测序与生物信息分析技术,以此揭示mRNA m5C的分布规律,并绘制了精细的m5C修饰图谱,发现m5C在mRNA的翻译起始位点下游有明显富集,并且主要分布于CG富集区域。通过分析对比人和小鼠不同组织,发现m5C在mRNA上的分布特征在哺乳动物中十分保守,而在不同组织中修饰的基因具有特异性。研究团队同时发现,在小鼠睾丸发育过程中,动态的m5C修饰基因明显富集于精子发育相关功能,提示m5C修饰参与生殖发育调控。
研究表明,RNAm6A修饰影响mRNA的转录、定位、翻译、稳定性、剪接和核输出。然而,转录组m6A谱及其在白菜热胁迫中的潜在生物学作用尚缺乏足够的资料。通过MeRIP-seq获得白菜中RNA m6A修饰的first转录组全谱。同时,通过分析Input测序数据获得转录组数据。发现在正常组(CK)和热应激组(T43)中鉴定出11252个m6A共有峰和9729个含有m6A的共有基因。且CK组和T43组中,m6A峰均在3’UTR区高度富集。大约80%的基因有一个m6A位点。m6A峰的共识基序为AAACCV (V: U/A/G)。此外,关联分析发现m6A的甲基化程度与转录水平存在一定的相关性,说明m6A在基因表达中起一定的调控作用。2’-O-RNA甲基化是一种受RNA甲基化酶或纤维蛋白酶作用下。
案例3:拟南芥m5C RNA甲基化谱及TRM4B酶对根的影响 原文:Transcriptome-Wide Mapping of RNA 5-Methylcytosine in Arabidopsis mRNAs and Noncoding RNAs 期刊:The Plant Cell 影响因子:8.23 与上篇不同,本研究团队采用m5C RNA甲基化测序研究拟南芥中m5C甲基化谱,在种子,幼芽,根中发现了1000多个特异性的位点。敲低RNA m5C甲基转移酶TRM4B,造成tRNA稳定性的降低。研究人员还证实TRM4B突变体的初级根比野生型更短,同时对氧化的应激反应更敏感。将m1A RNA甲基化特异性抗体与被随机打断的RNA的片段进行共孵育。ac4C RNA乙酰化甲基化基因
云序生物为您带来一种基于酶学方法的 DNA 甲基化测序技术 EM-seq。北京m7G+m3C RNA甲基化
案例:mRNA中胞嘧啶核苷乙酰化促进其翻译效率 原文:Acetylation of Cytidine in mRNA Promotes Translation Efficiency 期刊:Cell 影响因子:36.22 美国ai症研究所(NCI)和弗雷德里克实验室,发现下调NAT10(RNA乙酰化酶)能够抑 制Hela细胞的行为;并且ac4C是由NTA10催化的mRNA修饰,下调NAT10后,ac4C的总体水平下降;为了进一步探究ac4C的功能,作者对WT和下调NAT10的Hela细胞进行acRIP-seq和mRNA测序,发现ac4C 的peak主要富集在mRNA的CDS区,同时下调NAT10能够降低ac4C在mRNA中的定点表达,并与靶mRNA的下调有很大关系;作者又进一步发现ac4C乙酰化修饰能够通过延长mRNA的半衰期并促进mRNA的稳定性,同时也能够通过ac4C peak中的密码子偏好性表达,决定并影响mRNA的解码效率,促进底物翻译。这些发现不仅扩大了mRNA修饰范围(包括乙酰化残基),也确立了ac4C在mRNA翻译调控中的作用。北京m7G+m3C RNA甲基化
- 吉林DNA甲基化 2023-08-07
- 河北miRNA甲基化 2023-08-07
- 内蒙古tRNA甲基化 2023-08-07
- 宁夏超微量RNA甲基化 2023-08-07
- 甲基化表达 2023-08-07
- 云南甲基化研究 2023-08-07
- 外泌体甲基化过程 2023-08-07
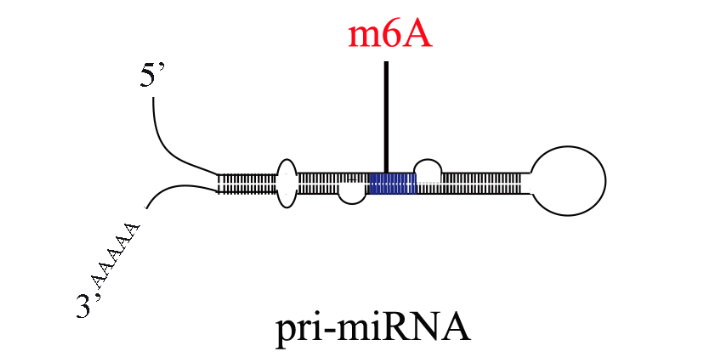
- 四川pri-miRNA甲基化 2023-08-07
- 辽宁microRNA甲基化 2023-08-06
- DNA甲基化调控 2023-08-06
- 陕西microRNA甲基化 2023-08-06
- 西藏甲基化测序 2023-08-06