- 品牌
- 上海云序生物
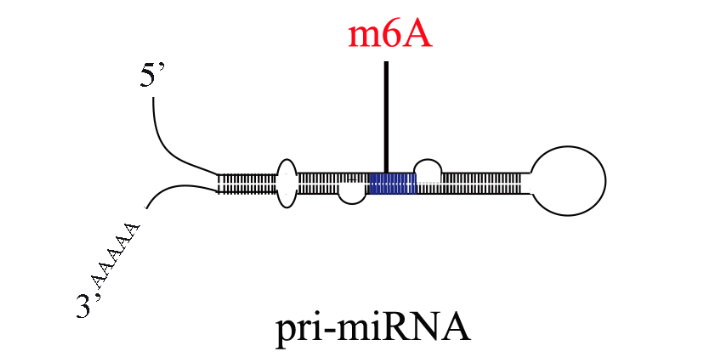
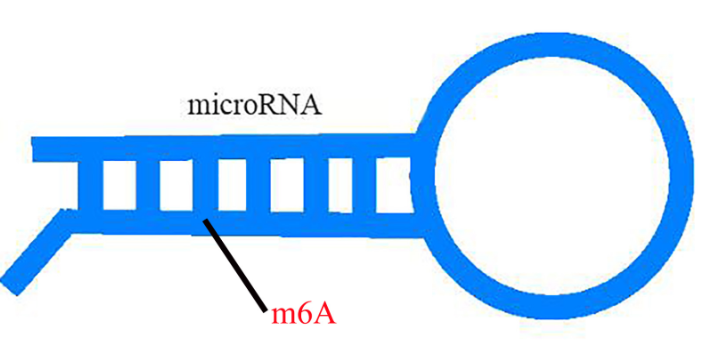
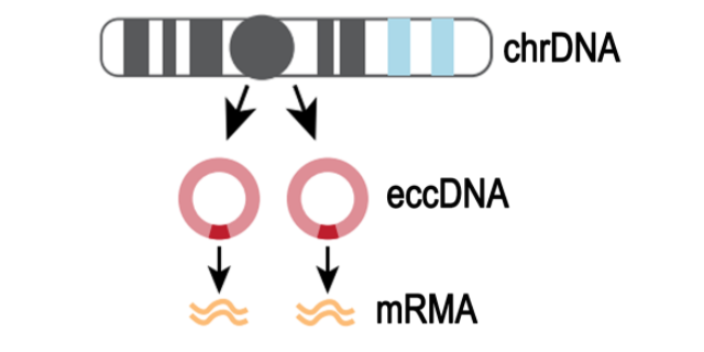
m6A甲基化已经被证明与植物对病原体的抗性有关。然而,小麦(Triticum aestivum) 全转录组m6A谱及其在小麦抗小麦黄花叶病毒(WYMV)中的潜在生物学功能尚未见报道。这项研究是shou次鉴定两个不同抗WYMV小麦品种的转录组m6A修饰谱。通过对m6A-MeRIP-seq数据的分析,作者在WYMV感ran的抗病小麦品种(WRV)和WYMV感ran的敏感小麦品种(WSV)中鉴定出25752个共有m6A峰和30582个共有m6A基因,这些峰主要富集在编码序列3‘UTR区和终止密码子中。GO分析和RNA测序数据显示,m6A和mRNA水平均发生明显变化的基因与植物防御反应有关。对m6A RNA甲基化,目前较流行的检测手段为m6A-Seq技术。m6A RNA甲基化
样本要求 样品类型 细胞、组织或RNA,其他样本类型请电询。 测序样品量 1.单碱基测序方式: a) 细胞:≥1×108 b) 组织:500 mg - 5 g c) RNA:100 μg - 300 μg 2. MeRIP测序方式: a) 细胞:≥2×107 b) 组织:100mg - 1g c) RNA:30 μg - 300 μg d) 超微量满足500 ng - 30 μg 样品运输及保存 样品运输:样品置于1.5mL离心管中,封口膜封好,干冰运输。 样品保存:细胞样品或新鲜组织块可用TRIZOL或RNA保护剂处理,液氮冻存后-80℃保存;RNA样品可溶于乙醇或RNA-free的超纯水中,-80℃保存,避免反复冻融。河南甲基化调控研究表明,m7G RNA甲基化修饰存在于各类分子中。
LncRNA m6Am-Exo-seq 测序服务 云序生物在国内shou批引入 m6Am-Exo-seq 测序服务,利用 5’ 核酸外切酶消除非目的性的 RNA 的片段上 m6A 修饰的信号干扰,选择性地获取 mRNA以及 LncRNA 的 5’ 帽子结构下游 m6Am 修饰富集区域的序列信息。接下来,我们对选择性获取到的 m6Am 修饰的 RNA 的片段进行反转录建库和高通量测序(测序结果将同时包含 mRNA 和 LncRNA)。通过后续的生物信息学分析,可以区分来自 mRNA 和 LncRNA 的测序片段,从而较全揭示 LncRNA和mRNA 的 m6Am 修饰位点,并推测其可能的生物学功能。
云序优势 单碱基分辨 m7G RNA甲基化单碱基测序方式,可对m7G甲基化修饰进行单碱基定位。 抗体富集效率高 m7G MeRIP测序方式,采用预验证的商业化抗体和精心优化的实验流程,具有极高的效率和特异性。 高通量检测 可对全转录组范围内的m7G位点进行高通量地平行检测。 全分子覆盖 地对环状RNA、mRNA、lncRNA、pri-miRNA、tRNA和rRNA等多类RNA分子的m7G位点进行检测。 一站式服务 客户需提供组织或细胞,云序生物一站式完成RNA抽提,样品预处理,建库,测序,数据分析流程。 专业的生物信息学分析 专业的生物信息学团队,能够满足客户的各类深入数据分析需求RNA甲基化组揭示了m6A介导的DNA去甲基化酶基因SlDML2在番茄果实成熟中的调控作用。
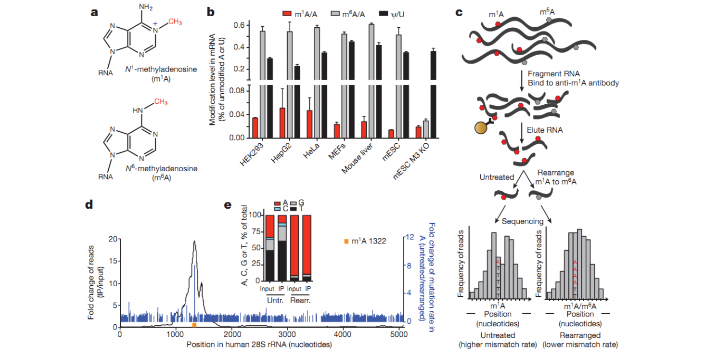
ac4C RNA乙酰化是在RNA ac4C修饰酶的作用下,使N4位乙酰胞嘧啶发生乙酰化的一种保守的化学修饰(N4-acetylcytidine)。早期研究发现该修饰存在于真核生物中丝氨酸及亮氨酸tRNA和18S rRNA上,导致Watson-Crick碱基热稳定性增加,调控蛋白合成中的编码准确性;近期研究发现ac4C分布在人类转录组中,大多数位点出现在编码序列(CDS)内,并且通过改善的mRNA稳定性和翻译促进靶基因表达。目前,ac4C RNA乙酰化修饰作为一类新型RNA修饰,是继m6A修饰之后的又一表观转录组学热点和“超级潜力股”!科学家们发现了一种可逆的RNA甲基化—m6A。四川甲基化机制
m6A甲基化已经被证明与植物对病原体的抗性有关。m6A RNA甲基化
研究还发现,m6Am 修饰是一个可逆的动态修饰,当细胞遭遇热激、低氧等应激性刺激时 m6Am 水平上升,说明 m6Am 可能在细胞应激反应中扮演了重要角色。近来也有研究发现,除了 mRNA 的较早编码核苷酸残基以外,在 snRNA 内部也有 m6Am 甲基化修饰的分布。虽然 m6Am 修饰早在 1975 年就已经被人类发现,但由于检验手段的匮乏,科学家难以区分出 m6A 修饰与 m6A 修饰,因此直到近年来 m6Am 测序技术逐渐发展成熟,才为进一步深入研究 m6Am 这一重要的 RNA 修饰铺平了道路。m6A RNA甲基化
- 吉林DNA甲基化 2023-08-07
- 河北miRNA甲基化 2023-08-07
- 内蒙古tRNA甲基化 2023-08-07
- 宁夏超微量RNA甲基化 2023-08-07
- 甲基化表达 2023-08-07
- 云南甲基化研究 2023-08-07
- 外泌体甲基化过程 2023-08-07
- 四川pri-miRNA甲基化 2023-08-07
- 辽宁microRNA甲基化 2023-08-06
- DNA甲基化调控 2023-08-06
- 陕西microRNA甲基化 2023-08-06
- 西藏甲基化测序 2023-08-06