- 品牌
- 上海云序生物
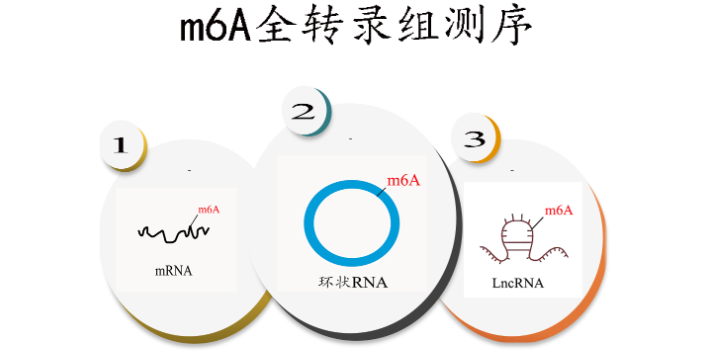
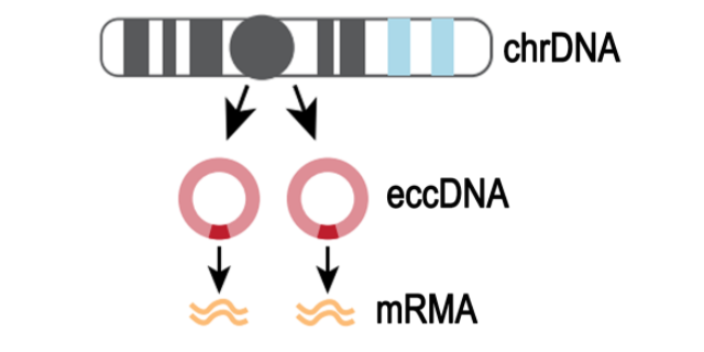
技术原理 为了系统性揭示RNA O8G氧化修饰,云序生物提供了成熟的 RNA O8G氧化MeRIP-Seq技术服务。技术原理如下:将RNA O8G氧化特异性抗体与被随机打断的RNA的片段进行共孵育,抓取有O8G氧化修饰的片段进行测序;同时需要平行测序一个对照(Input)样本,对照样本为未进行IP反应的RNA的片段。对照样本用于消除非特异性抓取O8G氧化片段的背景。对比免疫共沉淀IP样本和Input样本中的序列片段,将O8G氧化修饰位点定位到转录组上,并根据RNA-seq数据,计算样本中O8G氧化程度。 配合专业的生物信息学分析,云序生物可进一步提供高精度的O8G氧化图谱,帮助客户揭示O8G氧化在生物学功能和潜在的作用机制。种种研究迹象表明m6A 存在于很多物种中,占到了 RNA甲基化修饰的80%。浙江m7G RNA甲基化
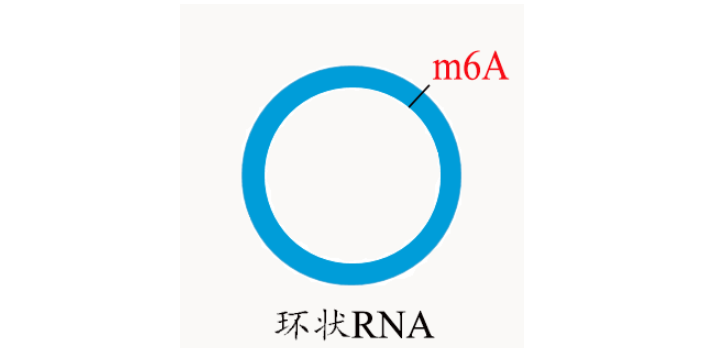
案例3:拟南芥m5C RNA甲基化谱及TRM4B酶对根的影响 原文:Transcriptome-Wide Mapping of RNA 5-Methylcytosine in Arabidopsis mRNAs and Noncoding RNAs 期刊:The Plant Cell 影响因子:8.23 与上篇不同,本研究团队采用m5C RNA甲基化测序研究拟南芥中m5C甲基化谱,在种子,幼芽,根中发现了1000多个特异性的位点。敲低RNA m5C甲基转移酶TRM4B,造成tRNA稳定性的降低。研究人员还证实TRM4B突变体的初级根比野生型更短,同时对氧化的应激反应更敏感。黑龙江m1A RNA甲基化环状RNA也会被m6A甲基化修饰,并会促进环状RNA编码蛋白这一有趣的结论。
m6A是真核生物中较常见的RNA修饰,被认为是一种新的表观遗传标记,参与多种生物过程。m6A调控几种主要人类病毒性疾病的模式和功能已经有报道。然而,m6A在植物病害暴发中的分布模式和作用尚不清楚。在此,作者分析了两种破坏性的病毒感ran水稻植株中的m6A修饰情况。发现m6A甲基化主要与病毒gan染水稻植株中表达不活跃的基因有关。作者还检测到同一基因上不同的m6A峰分布,这可能是水稻条纹病毒和水稻黑条纹矮病毒感ran之间存在不同抗病毒模式的原因。有趣的是,病毒感ran后水稻m6A甲基化水平增加。
研究还发现,m6Am 修饰是一个可逆的动态修饰,当细胞遭遇热激、低氧等应激性刺激时 m6Am 水平上升,说明 m6Am 可能在细胞应激反应中扮演了重要角色。近来也有研究发现,除了 mRNA 的较早编码核苷酸残基以外,在 snRNA 内部也有 m6Am 甲基化修饰的分布。虽然 m6Am 修饰早在 1975 年就已经被人类发现,但由于检验手段的匮乏,科学家难以区分出 m6A 修饰与 m6A 修饰,因此直到近年来 m6Am 测序技术逐渐发展成熟,才为进一步深入研究 m6Am 这一重要的 RNA 修饰铺平了道路。miRNA的生物合成和生物学功能、tRNA稳定性、18S rRNA的核内加工及成熟。
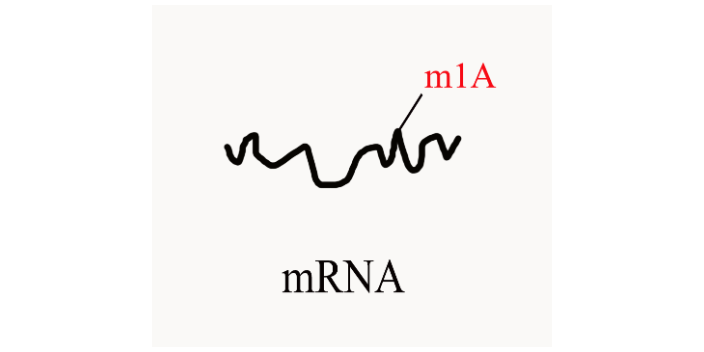
原文:METTL4 catalyzes m6Am methylation in U2 snRNA to regulate pre-mRNA splicing 期刊:Necleic Acids Research 影响因子:16.48 新加坡南洋理工大学的研究人员通过人类全转录组 RNA 甲基化测序,在 Mettl4 敲除组与野生型对照组之间寻找出 m6Am 相对甲基化水平的差异位点,其中 U2 snRNA 的第 30 位 RNA 残基有明显的 m6Am 修饰水平差异。进一步的 FLAG-tag 融合蛋白实验证明,METTL4 蛋白直接催化了 U2 snRNA 上的 m6Am 甲基化修饰的发生。METTL4 过表达实验发现,其催化的 RNA 内部 m6Am 修饰位点具有 HMAGKD 的序列 motif 特征(H=A/C/U, M=A/C, K=G/U, D=A/G/U)。 在 Mettl4 敲除的人类细胞中,因为 U2 snRNA 无法完成 m6Am 修饰,故而对 snRNA 的 pre-mRNA 剪接功能产生了诸多影响。2’-O-RNA甲基化修饰在mRNA、tRNA、rRNA、miRNA等分子上分布。陕西m7G RNA甲基化
m7G RNA甲基化单碱基测序方式,可对m7G甲基化修饰进行单碱基定位。浙江m7G RNA甲基化
案例1:哺乳动物mRNA内m7G甲基化转录组图谱 期刊:Molecular Cell 影响因子:14.25 由于全部种类的反转录酶均无法将RNA m7G位点逆转录成对应发生碱基突变的cDNA,为了准确的探究RNA内m7G甲基化情况,何川团队利用m7G自带正电荷的特征,开发了新的m7G单碱基深度测序方法。该方法能够将RNA内含m7G位点转化为另一种可产生反转录碱基变异的新位点,并依据碱基变异率估计m7G位点的甲基化水平。此方法随后也被证实可检测18S rRNA中的1639位内含m7G位点以及可揭示人类细胞tRNA中的22个46位内含m7G位点。并观测到其在mRNA分布、富集的共有序列及其它统计特征与m7G-MeRIP-seq数据基本保持一致。该文章不仅揭示了m7G甲基化在人类细胞中的分布特征,同时还发现METTL1是一种甲基转移酶,它在mRNA中催化了m7G甲基化修饰,并表明m7G的内部甲基化可以影响mRNA的翻译。浙江m7G RNA甲基化
- 吉林DNA甲基化 2023-08-07
- 河北miRNA甲基化 2023-08-07
- 内蒙古tRNA甲基化 2023-08-07
- 宁夏超微量RNA甲基化 2023-08-07
- 甲基化表达 2023-08-07
- 云南甲基化研究 2023-08-07
- 外泌体甲基化过程 2023-08-07
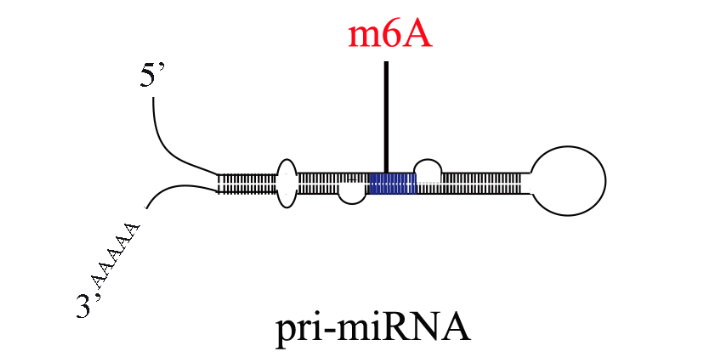
- 四川pri-miRNA甲基化 2023-08-07
- 辽宁microRNA甲基化 2023-08-06
- DNA甲基化调控 2023-08-06
- 陕西microRNA甲基化 2023-08-06
- 西藏甲基化测序 2023-08-06