- 品牌
- 上海云序生物
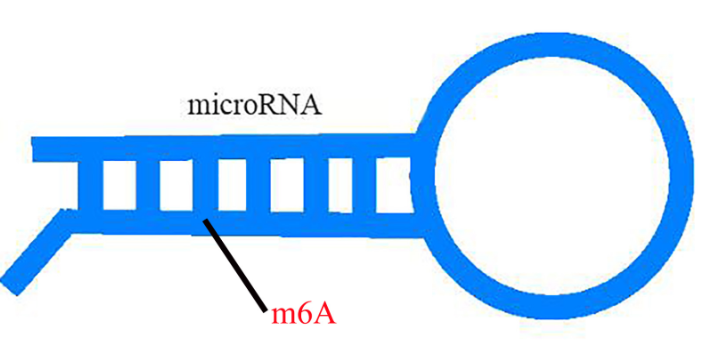
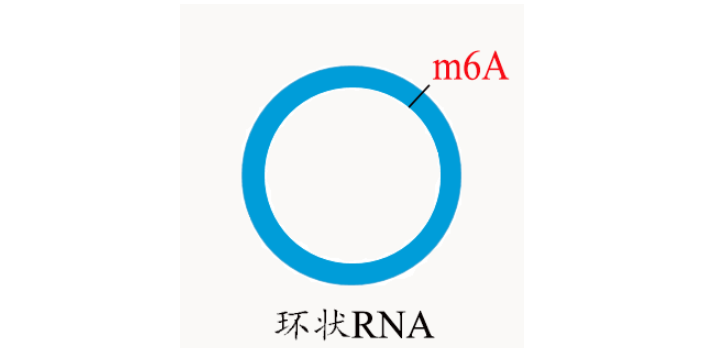
云序优势 单碱基分辨 m7G RNA甲基化单碱基测序方式,可对m7G甲基化修饰进行单碱基定位。 抗体富集效率高 m7G MeRIP测序方式,采用预验证的商业化抗体和精心优化的实验流程,具有极高的效率和特异性。 高通量检测 可对全转录组范围内的m7G位点进行高通量地平行检测。 全分子覆盖 地对环状RNA、mRNA、lncRNA、pri-miRNA、tRNA和rRNA等多类RNA分子的m7G位点进行检测。 一站式服务 客户需提供组织或细胞,云序生物一站式完成RNA抽提,样品预处理,建库,测序,数据分析流程。 专业的生物信息学分析 专业的生物信息学团队,能够满足客户的各类深入数据分析需求对m6A RNA甲基化,目前较流行的检测手段为m6A-Seq技术。江西甲基化表达
样本要求 样品类型 细胞、组织或RNA,其他样本类型请电询。 测序样品量 1.单碱基测序方式: a) 细胞:≥1×108 b) 组织:500 mg - 5 g c) RNA:100 μg - 300 μg 2. MeRIP测序方式: a) 细胞:≥2×107 b) 组织:100mg - 1g c) RNA:30 μg - 300 μg d) 超微量满足500 ng - 30 μg 样品运输及保存 样品运输:样品置于1.5mL离心管中,封口膜封好,干冰运输。 样品保存:细胞样品或新鲜组织块可用TRIZOL或RNA保护剂处理,液氮冻存后-80℃保存;RNA样品可溶于乙醇或RNA-free的超纯水中,-80℃保存,避免反复冻融。新疆microRNA甲基化m6A修饰在各种真核生物、各类组织细胞中普遍存在。
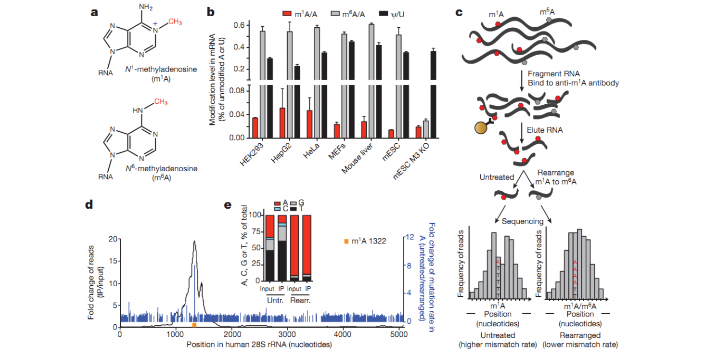
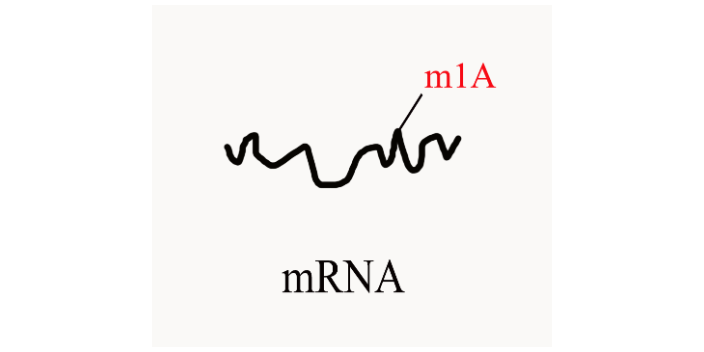
m1A RNA甲基化是一类新发现的RNA甲基化,即RNA分子腺嘌呤第1位氮原子上的甲基化修饰(N1-methyladenosine,m1A)。研究表明,m1A是真核生物tRNA和rRNA丰度很高的一种转录后修饰,近期研究也表明m1A修饰可调控mRNA翻译。m1A修饰作为一类新型RNA甲基化,其功能和机制都亟待挖掘。 与m6A RNA检测方式一致,针对m1A修饰也采用MeRIP-seq技术,抗体富集的技术原理如下:将m1A RNA甲基化特异性抗体与被随机打断的RNA的片段进行共孵育,抓取有甲基化修饰的片段进行测序;同时需要平行测序一个对照(Input)样本,对照样本为未进行IP反应的RAN片段,对照样本用于消除非特异性抓取甲基化片段的背景。
Nature | miR-1的位置特异性氧化导致心脏肥大 发表时间:2020年8月5日 影响因子:42.778 Nature上发表的来自韩国高丽大学Sung Wook团队的一篇研究在氧化还原相关疾病心肌肥大的大鼠模型中对氧化的miRNA进行了特异性测序研究。 作者在氧化还原相关疾病心肌肥大的大鼠模型中对氧化的microRNA(miRNA)进行了特异性测序。结果发现8-氧代鸟嘌呤2(o8G)修饰是选择性地在miRNA的特殊区域(位置2-8)产生的,并通过o8G-A碱基互补配对来调节其他mRNA。用肾上腺素能激动剂医治后主要在miR-1(7o8G-miR-1)的第7位诱导了o8G氧化修饰。单独引入的7o8G-miR-1或7U-miR-1(其中7位的G被U取代)足以引起小鼠心脏肥大,o8G-miR-1的mRNA靶基因在受影响的表型中起作用,反之对小鼠心肌细胞7o8G-miR-1的特异性抑制可减轻心脏肥大。o8G-miR-1也与心肌病患者有关。本研究表明miRNA的位置特异性氧化可以作为表观转录机制来协调病理生理学氧化还原介导的基因表达,为研究氧化修饰在病理生理学中的作用提供了新的思路。环状RNA也会被m6A甲基化修饰,并会促进环状RNA编码蛋白这一有趣的结论。
在这篇文章中,发现番茄果实成熟过程中,mRNA m6A甲基化表现出与DNA甲基化相似的动态变化。RNA甲基组分析表明,m6A甲基化是番茄果实mRNA中普遍存在的修饰,且m6A位点富集于终止密码子周围和3'UTR区。在具有DNA超甲基化的成熟缺陷表观突变体无色非成熟(Cnr)的果实中,有1100多个转录本显示m6A水平升高,而只有134个转录本显示m6A水平降低,表明m6A的整体水平升高。m6A沉积通常与转录本丰度呈负相关。云序优势 一站式服务: 客户只需提供细胞、组织或RNA,云序生物为您完成从MeRIP富集,文库制备,上机测序到数据分析整套服务流程。 miRNA的生物合成和生物学功能、tRNA稳定性、18S rRNA的核内加工及成熟。陕西microRNA甲基化
RNA m6A甲基化是RNA较关键的内部修饰之一。江西甲基化表达
案例:mRNA中胞嘧啶核苷乙酰化促进其翻译效率 原文:Acetylation of Cytidine in mRNA Promotes Translation Efficiency 期刊:Cell 影响因子:36.22 美国ai症研究所(NCI)和弗雷德里克实验室,发现下调NAT10(RNA乙酰化酶)能够抑 制Hela细胞的行为;并且ac4C是由NTA10催化的mRNA修饰,下调NAT10后,ac4C的总体水平下降;为了进一步探究ac4C的功能,作者对WT和下调NAT10的Hela细胞进行acRIP-seq和mRNA测序,发现ac4C 的peak主要富集在mRNA的CDS区,同时下调NAT10能够降低ac4C在mRNA中的定点表达,并与靶mRNA的下调有很大关系;作者又进一步发现ac4C乙酰化修饰能够通过延长mRNA的半衰期并促进mRNA的稳定性,同时也能够通过ac4C peak中的密码子偏好性表达,决定并影响mRNA的解码效率,促进底物翻译。这些发现不仅扩大了mRNA修饰范围(包括乙酰化残基),也确立了ac4C在mRNA翻译调控中的作用。江西甲基化表达
- 吉林DNA甲基化 2023-08-07
- 河北miRNA甲基化 2023-08-07
- 内蒙古tRNA甲基化 2023-08-07
- 宁夏超微量RNA甲基化 2023-08-07
- 甲基化表达 2023-08-07
- 云南甲基化研究 2023-08-07
- 外泌体甲基化过程 2023-08-07
- 四川pri-miRNA甲基化 2023-08-07
- 辽宁microRNA甲基化 2023-08-06
- DNA甲基化调控 2023-08-06
- 陕西microRNA甲基化 2023-08-06
- 西藏甲基化测序 2023-08-06